Research
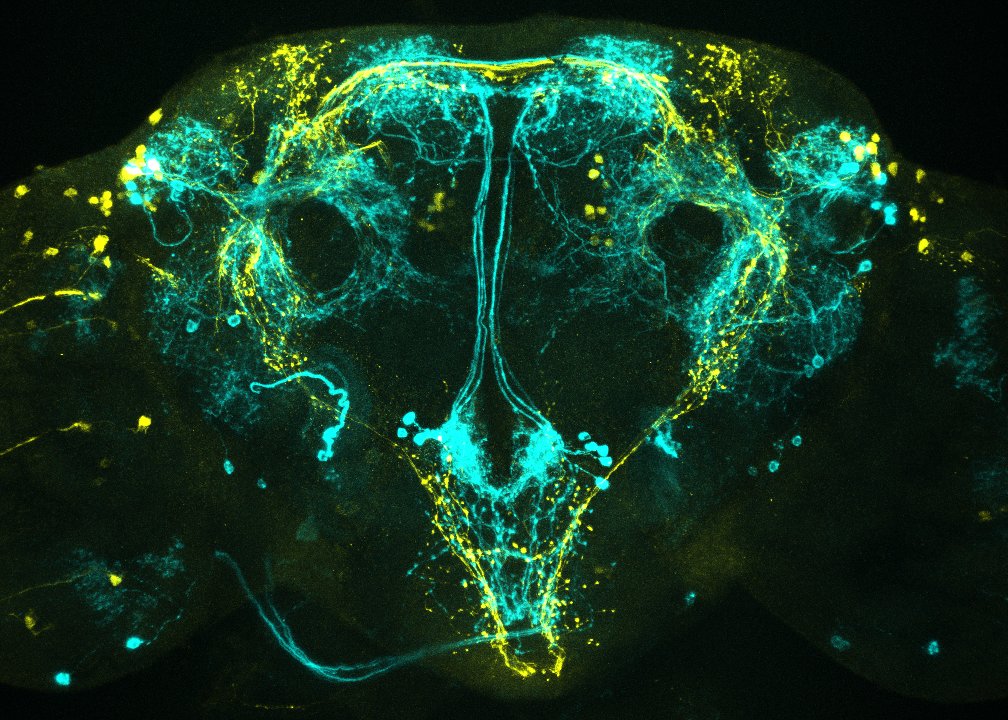
1. Identifying components of neural circuits that control social behaviors
A traditional view of social behaviors in animals is that they are stereotypical responses to sensory cues emitted by target individuals. While this relatively simplistic view has been useful in identifying key sensory cues and their receptors that guide social behaviors critical for survival (e.g., reproductive, parental, and aggressive behaviors) it is clearly not sufficient to account for the flexibility of these behaviors. One key element of social behavior is the ability of an animal to choose the correct behavior in response to what the other animal does. Even if each behavior is driven by a so-called “core circuit” consisting of a linear chain of neurons, information about the reactions of other animals and experiences from previous interactions must modulate the activity of the simple core circuit.
We have recently found that a dedicated set of neurons serve as a “brake” for aggressive interactions. These neurons release the neuromodulator, octopamine, which is a functional homolog of vertebrate norepinephrine. Taking advantage of the versatile genetic resources available in the fly, we are establishing genetic access to single octopaminergic neurons, enabling single-cell functional and physiological analyses. Using single-cell transcriptome analysis, we have identified several genes that are necessary to keep levels of aggression in check. Expression of these genes is orchestrated by an evolutionarily conserved transcriptional regulator that is present in these “brake” neurons.
We are also characterizing the functions of several neuropeptidergic cells that powerfully influence the levels of aggression in a context-dependent manner. We aim to uncover the neuronal components of an extended circuit governing aggressive interactions in the fly, which we will then use to perform genetic tests and decipher the computational principles of cost-benefit analyses.
2. The origins of sexually dimorphic social behaviors
Social behaviors of animals are often sexually dimorphic. Some behaviors are performed only by one sex, while other behaviors are performed differently between the two sexes. In flies, males perform species-specific courtship behaviors while females respond to male advances with either acceptance or rejection. Both male and female flies fight. However, males and females show distinct aggressive actions. Do these behavioral differences arise from differences in the circuits that control specific behaviors? Or do both sexes have the capability to perform all behaviors, but only certain behaviors are selectively activated in one sex?
Our neurogenetic analyses, as well as recent publications by other groups, argue that it is likely a little bit of both. While some sex-specific neurons are required to perform sex-specific behaviors, the presence of these neurons per se, or even forced activation of these neurons, is not necessarily sufficient to trigger the behavior. A likely scenario is that additional layers of neurons provide contextual information to allow animals to “release” a behavior only under proper circumstances. In the fly nervous system, these modules are often sexually dimorphic. Two sex-determining genes in the fly (doublesex and fruitless) coordinate specifications of distinct neuronal types in both courtship- and aggression-controlling circuits. Abrogating one of these two genes results in specific defects in the circuit and behavior, supporting the idea that sexual dimorphism in social behaviors results from the modular organization of the circuit.
3. Contexts that matter for social behaviors
What does “context-dependent” truly mean in social behavior? Researchers have noticed that the same sensory stimulus does not evoke the same behavior across individuals, or even within the same individual at different times. This type of variability is sometimes attributed to an “internal state”, an unobservable variable that controls the propensity of an animal to perform a particular behavior. Although a convenient word with an implication to “motivation” or “emotion”, an internal state is also a vaguely defined scientific term. What is the adaptive value of these states? We posit that “internal states” influence animal behavior by affecting the cost-benefit analysis of a given behavior. This is a particularly important calculation during conflict, in which both cost and benefit fluctuate moment-to-moment. We are currently focusing on two such factors: nutritional status and the experience of defeat.
An animal’s nutritional status can powerfully influence many behaviors. Hunger is an “internal state” that clearly functions to promote animal survival. It is unclear whether all types of “hunger” is the same, or if the deprivation of specific types of nutrition trigger specific state changes. In the context of social interactions, a limited food source promotes competition if animals are desperate to replenish their energy reserves. We found that flies are indeed capable of changing levels of aggression according to their nutritional states and the availability of resources. This phenomenon is nutrition-specific. Flies do not fight if they experience severe, general starvation, and flies deprived of one type of nutrition do not fight over another type of nutrition. Our preliminary data indicate that a specific neuropeptide is likely critical for converting the lack of a specific nutrient into a heightened level of aggression.
Social defeat is a type of learning. Once an animal is defeated in a conflict, it becomes less likely to engage in subsequent fights. Although often framed as a type of stress-related behavior, social defeat is an adaptive behavioral change if an animal does not have sufficient fighting capacity to successfully repel opponents. In other words, a defeat is not a loss per se. It is shown in wild populations that avoiding a fight can increase the fitness of a physically inferior individual. Social defeat in the fly is well documented. Using genetic access to specific aggression-controlling neurons, we are characterizing a specific circuit node that introduces information about social defeat. We aim to understand the phenomenon of social defeat not merely as an expression of stress, but as an adaptive behavioral change mediated by a dedicated circuit whose properties are defined by neurotransmitters and genes that define neuronal function.
4. Objective, reproducible, and quantitative measurements of social behaviors
Computational approaches for quantifying behaviors have emerged over the past decade, thanks to machine vision, machine learning (including artificial intelligence), and cheap and powerful computational resources. Although this is a welcome development, dense, multi-dimensional behavioral data are not necessarily accessible, interpretable, or even transparent. To be useful for neuroscience research, data generated by computational approaches should be biologically interpretable and, most importantly, reproducible among the community. Objective evaluation and data accessibility are key features of a quantitative behavioral dataset.
In a recent study, we evaluated the performance of computational behavioral classifiers under multiple behavioral conditions and quantified the impact of key parameters on the accuracy of behavioral quantification. It is safe to conclude that no computational approach is a “turn-key” system. Every algorithm has its biases and weaknesses, and it is the responsibility of each user to critically benchmark the performance of a computational approach. We deposited all of our movies, classifiers, and results of manual annotations on a public server so that everyone can validate our results for themselves. In an ongoing effort to establish unsupervised, comprehensive classifications of fly social interactions, we aim to prioritize biological interpretability and transparency. Our goal is to be a part of a community effort to establish common protocols for quantifying and sharing behavioral data. A universal platform for behavior quantification will maximize the positive impact of behavioral “big data” research.
5. Toward understanding the ultimate function of social behaviors
In a framework of social interactions, behaviors performed by others are as relevant as behaviors an animal performs to others. Social defeat is an example in which behaviors (aggression) performed by others has a notable impact on the function of the nervous system. In some animal species, social hierarchy (or dominant-submissive relationships) directly impact animal fitness by controlling reproductive success. We are interested in neural mechanisms by which certain social insect species (such as sweat bees and Harpegenathos ants) determine the reproductive (“queen”) caste through dominant-submissive relationship. While the effect of social defeat decays with time in the fly, hierarchy in these social insects has a lasting impact on their nervous system.
In humans, social isolation is considered a major risk factor in the context of longevity. We are currently investigating how social environments impact the fly lifespan. Previous studies have shown that group-rearing is generally detrimental to the lifespan of both male and female flies. Interestingly, we found that a co-housing with a specific fruitless mutant can extend (rather than shorten) the lifespan of male flies, suggesting that flies may exhibit different lifespans depending on the social environment. We are currently investigating the genetic and behavioral factors that underlie lifespan plasticity in specific social environments.
